使用经杀虫剂处理的蚊帐控制疟疾与蚊子媒介摄食行为的变化有关,包括早期和室外对人类的叮咬。表型可塑性和遗传性对这些行为转变的相对贡献尚不清楚。阐明这些转变背后的机制对于预测对病媒控制的影响至关重要。
采用一种新型便携式半场系统(PSFS)对坦桑尼亚疟疾病媒阿拉伯按蚊叮咬时间遗传力进行了实验测定。野生的。采用人工着陆捕获法每小时采集的阿拉伯滨猿猴,根据捕获时间分为咬食早(18:00-21:00)、咬食中(22:00-04:00)和咬食晚(05:00-07:00)3类,分别置于不同的笼中。然后给蚊子喂血供其产卵和形成第一代子代(F1)。每个咬食时间表型类别的F1代分别饲养,与母代同时采血。所得卵用于产生F2代,用于遗传力测定。通过释放F2 An来评估遗传力。将arabiensis放入PSFS中,记录它们在人类着陆捕获时的咬人时间,并将其与它们的F0祖母进行比较。
在PSFS试验中,F2后代(早18:00-21:00,中22:00-04:00或晚05:00-07:00)的咬人时间与其野生捕获的F0祖母的咬人时间呈显著正相关,估计遗传力为0.110 (95% CI 0.003, 0.208)。早咬F0的F2比中、晚咬F0的F2更倾向于早咬。同样,中晚咬食F0的F2的咬晚概率高于早咬食F0的F2。
尽管有一定的遗传力,但我们的研究结果表明,咬人时间的一些变化可归因于加性遗传变异。因此,选择蚊虫叮咬时间可以有效地发挥作用,强调需要针对早期和室外叮咬蚊虫的控制方法。
目前最重要的疟疾控制干预措施是驱虫蚊帐(ITNs)和室内滞留喷洒(IRS)[1,2]。自2000年以来,这些工具在非洲避免了6亿多例临床病例[1]。这些干预措施的成功源于它们能够利用非洲蚊子媒介的叮咬和休息行为的关键方面,包括它们在睡眠时间在室内叮咬人类并在喂食后在室内休息的倾向[3,4]。非洲最重要的疟疾病媒物种冈比亚按蚊(Anopheles gambiae)种群[4,5]和福氏按蚊(Anopheles funestus)[4,5]通常表现出这些行为[6,7]。尽管ITNs和IRS取得了成功,但它们的有效性正在被蚊子的适应性所削弱,蚊子对这些干预措施产生了抗性或逃避。最值得注意的是杀虫剂耐药性的广泛出现[8]。此外,越来越多的证据表明,非洲和其他地区的病媒行为发生了变化[9,10,11,12],这使得病媒减少了与ITNs和IRS的接触[7]。虽然对杀虫剂抗性的分子和遗传基础[13,14]及其对疟疾传播的影响已经进行了广泛的研究[8,15],但对蚊子叮咬行为适应的基础[16,17]及其对病媒控制的影响知之甚少[3,7]。
与ITNs和IRS相关的蚊子行为变化包括:提前离开喷洒过杀虫剂的房屋[18],在黎明或黄昏人们没有ITNs保护时增加户外叮咬[10,19,20,21],增加以牲畜而不是人为食[22,23]。媒介物种之间进行这种行为适应的能力可能有所不同。例如,历史上,安。据报道,东非的冈比亚疟蚊几乎完全以人为食[24]、在室内和深夜[4,6],而其兄弟物种阿拉伯按蚊(Anopheles arabiensis)则更灵活地以人和牛为食[25,26],在室内或室外[20,27],经常在傍晚和黎明[20,28]。随着ITNs的广泛应用,An的相对丰度。冈比亚与安哥拉的比较。Arabiensis在几个环境中急剧下降[29,30],因为该物种高度倾向于在室内和深夜进食。这种行为发生的时间与大多数人在室内和使用ITNs的时间一致,因此增加了An的风险。冈比亚人与杀虫剂有致命接触。在西非,有报告称,科鲁兹按蚊(Anopheles coluzzii)的叮咬倾向于傍晚或清晨发生变化,户外叮咬更多[31]。Funestus[11,19]。同样,西南太平洋地区实施IRS后,疟疾媒介法劳按蚊(Anopheles farauti)“早”(18:00-21:00 h)和室外叮咬的比例也有所增加[10,32]。还有证据表明,在实施ITN和IRS之后,非洲疟疾病媒的宿主选择从人转向了牛[22,23]。
预测蚊子行为变化对病媒控制的影响需要了解这些变化的潜在机制。目前尚不清楚行为转变是否反映了对ITNs/IRS选择的进化适应,或者是先前存在的表型可塑性的表现。这些可能性对控制有不同的含义。第一个假设被称为真正的行为抵抗,即行为特征是在对干预选择的反应中进化的。因此,行为抗性特征可以在种群中传播并固定下来[3]。第二种假设被称为行为弹性[9],即媒介物种总是能够表达可选择的咬人表型,这种可塑性仅在应对减少人类宿主可用性的环境变化时表现出来。在这里,当控制干预解除后,咬人行为可能迅速恢复到基线表型[16]。行为弹性可以定义对干预的即时行为反应的限制[3],而行为抗性意味着媒介可以越来越多地适应其叮咬表型,以避免室内干预。因此,可以通过使用它们来防止的暴露比例逐渐减少。这种机制在一定范围内可能比塑性产生更大的问题。虽然多年来已经观察到ITNs的使用与蚊子叮咬时间行为的改变有关,但对蚊子寻找宿主的遗传基础的调查相对有限。
本研究对非洲主要疟疾病媒蚊虫叮咬时间的遗传力进行了研究。在坦桑尼亚,利用一种新型便携式半田系统(PSFS)对arabiensis进行了实验研究。在控制条件下,将野生捕获的母鱼(F0)与其后代(第二代)的咬人时间表型进行比较。
所有实验均在Kilombero山谷内的Lupiro村(?8.38 S, 36.67 E)进行,该地区是坦桑尼亚东南部疟疾中高地方性传播地区[33]。目前,一个。Arabiensis是该地区最丰富的疟疾媒介物种[34]。本地区的咬人活动。Arabiensis种群可以早在黄昏时开始活动,在午夜前后达到高峰,在早晨达到较小的高峰[27]。大多数居民在户外待到晚上10点左右,然后回家睡觉;大多数报告使用itn[27]。所有蚊子行为分析都是在一个定制的半场系统中进行的,这里称为便携式半场系统(PSFS),临时安装在Lupiro村(详见附加文件1)。PSFS位于收集野生蚊子(亲代)以产生用于实验的后代的同一村庄。
在卢皮罗村,寻找主人的女性安。在夜间不同时段采用人工着陆捕集(HLC)采集阿拉比ensis。在HLC中,一名志愿者坐在椅子上,露出双腿,并吸入试图以其为食的蚊子。采集地点在4所房屋附近的居民区(附加文件2)。简言之,志愿者在18:00至07:00之间每小时采集一次蚊子。连续两晚(2015年7月14日和15日),每小时采集45分钟,休息15分钟。每小时收集的蚊子被放置在单独的纸杯中。每个实验夜结束后清晨逐小时采集,目测蚊属冈比亚按蚊(Anopheles gambiae sensu lato, s.l)[4,5],并根据捕获时间将其分为叮咬早期(18:00-21:00)、叮咬中期(22:00-04:00)和叮咬后期(05:00-07:00)3类,分别置于单独的笼中。咬人活动被分为三个不相等长度的离散类别,以对应于人们可能在室内并受到ITNs保护的时间(中期),或在室外且不受保护的时间(早期和晚期)。在245只雌性大熊猫中。捕获冈比亚蚊218只(咬早组71只,咬中组98只,咬晚组49只)。如附加文件2所述,为这F0雌性提供血粉用于产卵。121个母鼠(分别为43个、51个和27个,分别为早、中、晚)的卵经PCR鉴定为An。这些HLC野生捕获PCR证实了An。arabiensis雌性个体构成FO亲代。每个人的蛋都证实了安。Arabiensis母亲分开饲养,过渡到幼虫,从而到蛹期。根据每只雌鸟的咬人时间表型,从每只雌鸟的卵窝中孵化出来的蛹被放入三个笼子中的一个。卵-蛹发育率的差异导致设置了9个F1笼,每个叮咬时间表型3个。每个叮咬时间表型类别的F1代分别饲养,并在收集其母亲的同一时间段(早期,中期或后期)进行血液喂养,以产生F2代用于PSFS的分析(附加文件2)。由于从F0到F1的卵-蛹发育率的变化,这意味着不是所有的F2都是在同一天获得的,而是根据羽化率而变化。F2维持在PSFS内的环境条件下,以葡萄糖为饲料来源,直到释放日。实验选择F2代,而非F1代,以最大限度地提高可用于实验的蚊子数量。
通过释放F2 An来评估遗传力。将arabiensis放入PSFS中,记录它们在人类着陆捕获时的咬人时间,并将其与它们的F0祖母进行比较。在实验的每个晚上,300f2an。在晚上17:00 h同时将arabiensis(来自早、中、晚F0表型的100株)释放到PSFS中,但有一个试验除外,该试验中三种表型各只有50株F2可用(附加文件2,统计分析方法见下文)。释放的蚊子年龄从4天到10天不等,既有年轻蚊子,也有年老蚊子[35]。在释放前2小时,从笼中选择2只雌蚊,放入纸杯中,根据其祖母的叮咬时间表型(18:00-21:00、22:00-04:00和05:00-07:00)标记红色、黄色或蓝色荧光尘色。试验每天15:00 h进行选择和标记。17:00 h将蚊子释放到PSFS中,在HLC开始重新捕获之前,蚊子在PSFS内定位至少1小时,然后再进入宿主。志愿者于18:00进入PSFS,在18:00至07:00之间坐在椅子上开始HLC(附加文件2)。每小时进行45分钟的重新捕获,志愿者休息15分钟。在每次用hplc进行夜间行为试验结束时,用手电筒彻底搜查PSFS内的地板、屋顶和墙壁,使用背包吸引器收集任何剩余的蚊子(未被hplc捕获)。这是为了确保在开始另一个实验之前,在一个夜间实验中使用的所有蚊子都被清除。2015年8月13日至10月1日共20晚进行了检测。使用气象站测量和收集夜间温度(附加文件2)。在同一天晚上,在PSFS附近的两个当地房屋(约40米范围内)进行了额外的hplc收集,作为实验分析,以确认在PSFS中观察到的咬人活动的时间模式是否与野生种群一致。从晚上18:00开始,于次日早晨07:00结束,在当地房屋内外采集野蚊。
在对F0和F2的咬人时间进行遗传和关联检验之前,对PSFS上F2后代和野生安家鸡的夜间咬人情况进行了分析。比较了两种方法在相同夜晚的阿拉伯芥寻找寄主的情况。通过比较,可以评估PSFS内蚊子的叮咬特征是否代表了自然的叮咬活动。利用lme4 R软件包的glmer函数[36]拟合logit-二项广义线性混合效应模型(glmm),分别估算每个地点(室内、室外和半野外)在每个时间段(早、中、晚)被捕获的蚊虫比例及其95%置信区间,其中叮咬时间建模为二项响应(早vs中+晚;中vs早+晚;深夜vs早+中)和实验夜(20个重复)拟合为观察水平随机效应。使用McCulloch等人[37]的近似方法纠正了由于Jensen不等式导致的预测比例和95% ci的偏差。
咬人时间h2的狭义遗传力(描述可归因于基因加性效应变异的表型变异的一部分)估计为h2=2tF2-F0,其中tF2-F0是祖辈-后代(F2)咬人时间与祖辈(F0)咬人时间之间的相关性(详细方法和估计h2的R代码见附加文件3和4)。由于未知程度的同型交配在F1代,这h2估计预计是积极的偏见,但我们发现,当h2值低于0.5时,这种偏差可能是中等到低的(< 17%的相对偏差),而当h2值低于0.3时,这种偏差可以忽略不计(< 4%)(补充信息3)。使用MCMCglmm软件包,将F0和F2咬入时间建模为混合效应有序概率GLMM中的有序分类响应(早
除了估计咬伤时间的遗传力外,还检验了个体F2咬伤时间表型是否与F0咬伤时间表型相关。每个F2咬伤时间表型被建模为二元反应(早期vs中期+晚期;中vs早+晚;使用lme4r包的glmer函数在logit-二项式GLMM中的后期vs早期+中期)[36]。F0咬人时间拟合为类别固定效应,随机效应拟合为日期和观测水平随机效应。对于每个二项F2反应,使用似然比检验检验与F0咬人时间无关的零假设。用Wald检验F0次咬人F2咬人时间比例的两两差异。根据拟合的glmm计算具有95%置信区间(CI)的预测F2咬人时间比例。使用McCulloch等人[37]的近似方法纠正了由于Jensen不等式[40]导致的预测比例和95% ci的偏差。
摘要
背景
方法
结果
讨论
结论
数据可用性
参考文献
致谢
作者信息
道德声明
补充信息
搜索
导航
#####
在20个晚上的采样中,从邻近PSFS的当地房屋收集了24,503只野生蚊子。其中28% (n=6883)为An。冈比亚s.l。经PCR扩增成功的冈比亚虫标本均为冈比亚虫。arabiensis。在5850f2安。arabiensis(起源于1290年亲本An)。在PSFS中释放的arabiensis进行咬人时间测定,82%被重新捕获(实验夜为52-98%)。
F2安的咬人时间模式。PSFS内的arabiensis在相同的夜晚与野生种群中观察到的相似(图1,表1);确认在PSFS中保持了代表性的咬人行为。在所有蚊虫收集(室内、室外或PSFS)中,大约三分之一的叮咬发生在夜间早期,三分之二发生在夜间中期,只有十分之一发生在夜间。
图1
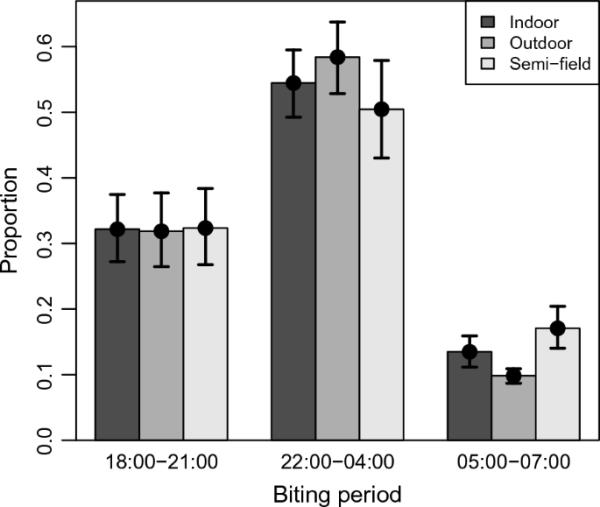
夜间不同时段阿拉伯按蚊叮人比例(早:18:00-21:00;中:22:00-04:00,晚:05:00-07:00)。条的颜色。误差柱为拟合模型的95 CI
表1 .的原始数据比较按蚊arabiensis在夜间不同时段被发现咬人
据估计,F2与F0的咬食时间表型之间存在0.055的弱正相关(tF2-F0 [95% CI]=0.055[0.001, 0.104])。利用h2=2tF2-F0的关系,这种相关性转化为估计的咬伤时间遗传力为0.110[0.003,0.208]。这种遗传力估计是足够低的,在估计中,由于分类交配的相对正偏差可能小于5%(附加文件3),因此不应影响结论,即大约十分之一的变异在An。阿拉伯人的咬人时间是由加性遗传变异引起的。
在每个二项glmm中,F2的咬入时间(定义为二元反应)和F0的咬入时间之间存在显著的正相关(表2)。早期和晚期咬入F2的咬入时间与其F0祖母的咬入时间呈正相关(图2,表2)。早期咬入F0的F2比中期咬入F0的F2更可能咬入早期(P < 0.001)。中期咬入F0的F2比晚咬入F0的F2更有可能早咬(P=0.018)(图2,表2)。在晚咬的概率上观察到类似的正相关,中期(P=0.029)和晚咬入F0的F2比早咬入F0的F2更有可能晚咬。然而,中咬和晚咬F0的后代在晚咬的概率上没有差异(P=0.29)。大多数F2咬人活动发生在“中期”(图2)。F2在F0早期和中期的咬人概率没有差异(P=0.066), F2在F0中期和晚期的咬人概率没有差异(P=0.19)。然而,晚咬的F0的F2较早咬的F0更有可能在中期咬伤(P=0.002,图2,表2)。
表2比例与95% coF2只蚊子在夜间早、中、晚3个时段叮咬的置信区间(95% CI),分别由F0只蚊子祖母的叮咬时间估算
图2
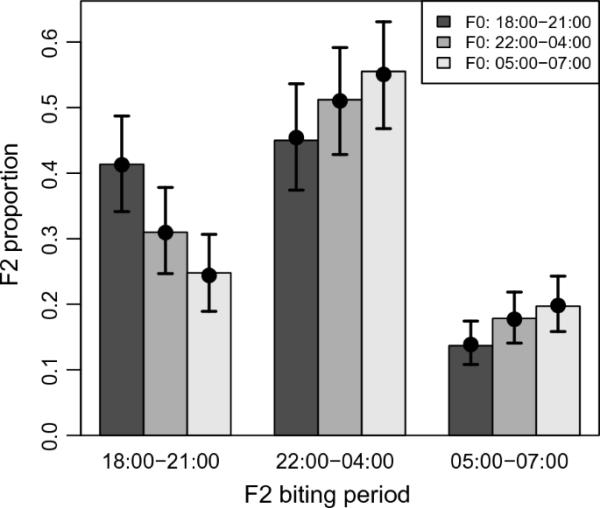
预测夜间不同时段雏鸟咬人比例(早:18:00-21:00 h)、中:22:00-04:00 h、晚:05:00-07:00 h)。柱状线颜色表示雏鸟咬人时间。误差柱为拟合模型的95% CI
本研究对非洲疟疾病媒An的叮咬时间倾向的遗传基础进行了实验研究。通过比较野生F0种群及其F2后代在实际半田间条件下的表型。遗传率估计为0.11,这证明了咬人时间的一部分自然变异——尽管是少数——是由于加性遗传变异。大多数昆虫研究遗传率的属果蝇的遗传率估计在0到60%之间[41]。在这种情况下,这里获得的11%的估计值相对较低。然而,当与其他行为特征单独比较时(与倾向于具有更高遗传力的形态特征相反),这一估计处于0-20%观察范围的中心。特别是,早期咬过F0的祖母的后代比中期或晚期咬过F0的后代更有可能咬得早。同样,晚咬F0的F2后代比早咬F0的后代更有可能晚咬。而中、后期咬人F0的后代在这两个时期的咬人频率相似。因此,在任何极端的咬食活动范围内(晚上10点之前或凌晨5点之后)F2进食的可能性与它们的亲代表型有关。总的来说,尽管估计遗传率相对较低(11%),但这表明选择可以影响野生非洲疟疾病媒的叮咬时间。
这一证据表明,后代和祖父母的“极端”咬人时间表型(例如:早:18:00-21:00 h,或晚:05:00-0.7:00)之间存在遗传关联,这表明在典型睡眠时间(22:00-05:00 h)使用ITNs可能会选择改变印度人的咬人时间。Arabiensis到大多数人没有保护的时期。这里故意选择将蚊子叮咬的时间划分为不等长的时间段;早期(4小时)、中期(6小时)和晚期(3小时),重点分析itn规划中特别存在问题的行为的遗传性。“中期”的延长可能解释了为什么大多数安。在这段时间内,无论其祖父母的咬人时间表型如何,都观察到阿拉伯人的咬人行为。如果人们在睡眠时持续使用ITNs,中期蚊虫叮咬时间的微小变化可能不会产生流行病学后果;通常发生在非洲社区的22:00-04:00之间[27,42]。相反,将蚊虫叮咬时间转移到人们进入室内睡觉前(晚上10点之前)或早晨醒来后(凌晨5点),将减弱ITNs的影响[7]。例如,最近来自西非的证据发现了安。由于ITNs的广泛使用,在人们外出的早晨(上午7点至11点),funestus咬人[11,19]。这一研究表明,安。在这些极端范围内咬人的阿拉伯人确实有遗传成分;这引起了人们的关注,即疟疾病媒可能会越来越多地根据杀虫剂或其他来源的选择来调整其叮咬时间。除了更广泛记录的生理杀虫剂抗性性状之外,这种行为适应性代表了疟疾病媒对干预措施产生抗性的另一种潜在机制。通过选择叮咬时间产生的行为抗性也可能导致传播的残留[7]和反弹[3]。
先前的一项研究也报道了亚马逊西部早期和晚期咬人Nyssorhynchus (Anopheles) darlingi遗传结构的证据[43]。然而,先前对早咬和晚咬的遗传变异的调查。本研究区域的阿拉伯人在一系列候选昼夜节律基因和单核苷酸多态性中未发现亚结构的证据[44]。这些作者承认,由于样本量和分析的snp数量的限制,该研究在咬人时间内检测遗传变异的能力较低[44];并且需要一个更强大的关联映射分析来得出结论。结果表明,黄杨的F0和F2的咬合力较低,但不为零。arabiensis表明,最近观察到的疟疾媒介叮咬时间对ITNs反应的变化[12,45]很可能是除了近乎瞬时的表型可塑性外,还源于扩展的进化过程。
这项研究可能有一些局限性,可能有偏见,可能低估了遗传性。最初的担忧是,在半野外条件下观察到的蚊子叮咬时间可能不能反映自然表型。然而,该研究证实,PSFS的夜间咬人活动模式与当地野生种群在相同夜晚观察到的相似。其次,在咬人时间内检测遗传的能力可能已经降低,因为不是比较母亲-后代对;分析是基于在每个时间段内由F0叮咬队列产生的汇总后代。为了产生足够大的行为生物测定样本量,这种设计是必要的,但在分析中要控制的单个蚊子母亲-后代对之间的差异无法识别。因此,F2咬人时间表型代表了一组祖母的平均表型,可能掩盖了特定父母-后代对之间更强的关联。第三,虽然只有121个F0的样本量可能有问题,但在本研究中是相当充足的。尽管遗传力估计较低,但根据补充信息(SI 3)中描述的低P值,FO-F2相关性是可靠的。值得关注的是,咬人时间被记录为三个类别,而不是连续变量。这些可能会限制估计的精度和能力。第四,在与雌蚊捕获时间相对应的时间给雌蚊供血F1可能使雌蚊产生条件反射,从而产生非遗传的母体效应[46]。虽然不能排除母体效应的存在,但在蚊虫宿主偏好表型的相关试点工作中未发现母体效应的证据。arabiensis (Govella et al.未发表),也不知道有任何其他证据表明母体效应会影响蚊子的叮咬时间。第五,在20个晚上的实验中,被释放的蚊子的年龄在4到10天之间。有可能由于与年龄相关的咬人变化而引入了一些变异。然而,先前对同一物种的研究并未发现咬人时间随年龄的变化[47]。最后,研究是在单个村庄和单个安进行的。Arabiensis种群数量,因此限制了结果的推广。尽管存在这些限制,但在咬人时间上发现了跨代相关性的有力证据。预计在这些设计上的潜在限制可以解决的条件下,进一步的调查将产生更精确的遗传性估计。该研究特别强调了在具有不同叮咬时间表型的蚊子种群之间进行交叉实验的价值,以确定结果是否支持孟德尔遗传模式。尽管本研究获得的遗传率较低,但这将为咬时表型的遗传提供证据。
这项研究提供了第一个实验确认的存在和遗传的程度,在咬时间的主要非洲疟疾媒介。这些结果为进一步研究这种表型和其他蚊子摄食行为(宿主偏好和叮咬地点)的遗传基础奠定了基础,这些行为在干预措施下的进化可能会破坏非洲消除疟疾的努力。这些结果表明,尽管安。阿拉伯人的咬人时间可能主要由非遗传因素驱动,这种行为表型也可能部分遗传。因此,在野外种群中观察到的叮咬时间变化可能受到干预引起的长期选择的影响,以及非遗传表型可塑性可能发挥的更大作用。无论这些过程的相对贡献如何,越来越多的证据表明,疟疾病媒种群的叮咬时间发生了变化,这突出表明迫切需要采取补充性干预措施,以室外环境和典型睡眠时间以外的蚊子为目标。
便携式半场系统。
咬合力测定的方案和实验设计。
咬伤时间遗传力估计的统计方法。
r码显示选型交配如何影响F1配偶之间的表型相关性以及由此产生的遗传力估计的预期偏差。
下载原文档:https://link.springer.com/content/pdf/10.1186/s12936-023-04671-7.pdf
为您推荐:
- 欧盟国家威胁要切断乌克兰的电力 2025-02-16
- 拉塞尔·布兰德称“骇人听闻”的强奸、殴打和精神虐待指控“非常伤人” 2025-02-16
- HIF-1活性直接导致乳腺癌扩散到淋巴管 2025-02-16
- 一名男子因涉嫌在克鲁克屋酒吧现场骚乱后袭击而被捕 2025-02-02
- 伯明翰糖果公司关闭皮兹食品大厅的位置 2025-02-02
- 我来教大家“边锋跑得快开挂教程方法!详细开挂教程已更新-知乎 2025-02-02


